
What is Sex?
What you didn't know you didn't know...

I initially wrote this post to help explain the concept of sex as used by clinicians in order to demystify disagreements around the percentage of intersex people and the degree to which sex is binary. However, after I posted my sex and gender dialogue Steersman alerted me to why the gametic definition of sex is primary and how the clinical definition is dependent on the gametic definition.
While the biological definition of sex may be primary, the clinical definition of sex is more relevant for some medical purposes and is much closer to the “folk” understanding of sex categories. The clinical definition also has the advantage of being able to classify a much larger percentage of people vs. the gametic definition.
Just as there are some areas where gender is the relevant consideration and some where sex is, there are also some areas where the biological definition of sex is relevant (for humans) and some where the clinical definition is. So with that…
This post has two parts:
Explaining the cross-species definition of sex from evolutionary biology
Explaining the clinical definition of sex and illustrating how it forms a tightly bimodal distribution rather than a spectrum or a strict binary
1. Sex as defined in Evolutionary Biology:
Source: Paul E. Griffiths, What are biological sexes?, University of Sydney (2021):
The understanding of sex in evolutionary biology serves to “address basic questions such as ‘why does sex exist?’, ‘why do some species have separate sexes?’, and ‘why do some species have sex chromosomes?’” (p. 7) in a way that is relevant across sexually reproducing species.
“In sexual species reproduction occurs when two gametes fuse to form a zygote. Gametes are the sex cells which contain all or part of the DNA of one parent.” (p. 2). In evolutionary biology the sex of an individual is defined based on the size of the gametes they produce: large gamete producers are female, small gamete producers are male and individuals which produce both types of gametes are hermaphrodites. In some species individuals are hermaphrodites simultaneously while in others they’re sequential hermaphrodites, switching from one sex to the other in their lifetime. Some species, such as bees, also have non-reproductive castes whose individuals cannot meaningfully be said to have a sex at any point.
Gamete size is the differentiating factor for evolutionary biologists because it describes the two basic reproductive strategies seen in all complex multi-cellular sexually reproducing species. These strategies involve organisms which either “produce many small gametes, making it more likely that some of them will find a partner, or produce fewer but larger gametes, making it more likely that the new individual will have what it needs to survive and thrive.” (p. 8).
While it’s possible for a species to evolve to produce more than two distinct sizes of gametes or gametes that vary in size continuously, due to competitive dynamics these forms of reproduction are not seen in practice1 with the exception of “a group of species at the boundary between unicellular and multi-cellular life” (p. 8). So, in essentially all sexually reproducing species, gamete size is binary. The behaviors and phenotypes associated with males and females often fit general patterns that we observe across many species but depend on how these “two basic reproductive strategies play out in different ecological conditions” (p. 10).
Sex chromosomes, which are key for sex determination in humans, cannot be used to define sex in general because many animals have males and females but do not have sex chromosomes2. Moreover, in humans and other animals that have chromosomal sex determination, “Biologists know which chromosome pairs are male or female because they know which animals are male or female, using the gametic definition” (p. 5), not the other way around. In other words, biologists know which humans are female vs. male based on which humans produce eggs vs. sperm. They then connect XX chromosomes to females and XY chromosomes to males by observing their relationship to sex determination in those gametically categorized populations.
However, “The claim that a species has only two biological sexes is commonly misunderstood as the claim that there are only two kinds of individual in that species.” (p. 5). A species can have different mating types3 within a sex, where certain types of males can only mate with certain types of females, or different morphs4 of that sex, where individuals of the same sex develop distinct phenotypes suited to different mating strategies. But, biologists find it useful to distinguish between sex and other phenomena such as mating types given that they evolved for different reasons and given the unique explanatory role that sex plays in “the later evolution of sex chromosomes, sex-associated physical characteristics and much more.” (p. 11).
As useful as this definition is for biologists however, it isn’t totally satisfactory when talking about how to medically classify humans because many humans (most obviously children and post-menopausal women) do not produce gametes and so are sexless by this definition.
“In a sense all complex multicellular organisms ‘change sex’ because they do not have a sex at conception and must develop one as they grow to reproductive maturity. [...] When we assign a sex to an organism not yet reproductively competent we are engaging in prospective narration.” (p. 17)
Thus, instead of viewing sex as an identity characteristic, this definition treats sex as a life-history stage, and “There is no biological reason why these individuals should have a determinate sex at every life stage or indeed at any life stage.“ (p. 21)
So, while “The biological understanding of sexes is an effective tool for studying the diversity of reproductive systems across species. It has not been developed to assign every individual organism to a sex and it often fails to do that...” (p. 15). In addition “Sex and sexes as aspects of human life and human society are about much more than the production of offspring. That is a good reason to reject an exclusively biological understanding of sex in the human and social sciences.” (p. 6). So, let’s move on to the (probably more familiar) clinical definition of sex.
2. Clinical Definition of Sex in Humans:
The clinical concept of sex is a biological construct with sex categories defined based on the presence of certain biological characteristics. My use of the word construct is not to imply that sex isn’t real and important or to deny that sex categories have been stable over time, but to illustrate that to define any category or concept, we have to agree on parameters that sequester one category from another.
So… what are the characteristics that are (generally) used to define sex categories medically and how do we categorize individuals as either male or female?
From the NIH, sex is a biological construct defined based on:
Genotypic sex (generally XX or XY chromosome)
Phenotypic sex (observable sex characteristics/how your body looks and functions)
Anatomy:
Internal/external genitalia (e.g. prostate, penis etc. or uterus, vagina etc.)
Secondary sexual characteristics (e.g. boobies)
Physiology
Hormones
Most people with the XX genotype are phenotypically female and most people with the XY genotype are phenotypically male. We use the term intersex for people who don’t fit that typical pattern. The type of gametes produced (if any) are also part of an individual's phenotypic expression but are not specifically referred to in the clinical definitions of sex that I found.
Is Sex Binary or Is Sex a Spectrum?
You could imagine creating a sex score. For example:
Sex Score = - 5*(Has a Y chromosome) + 5*(Has no Y chromosome) - 4*(Has male genitalia) + 4*(Has female genitalia) - 1*(Has male secondary sexual characteristics, hormones and physiology) + 1*(Has female secondary sexual characteristics, hormones and physiology)
(Has a Y chromosome) and (Has no Y chromosome) are both binary variables (True/False or 1/0). For simplicity, let’s also pretend that (Has male genitalia) and (Has female genitalia) are binary. Finally, let’s allow (Has male secondary sexual characteristics, hormones and physiology) and (Has female secondary sexual characteristics, hormones and physiology) to be continuous variables which each average 1 across people who have a Y chromosome and people who have no Y chromosome respectively.
Then, if we scored a “typical female” and a “typical male” they would get the following scores:
Typical Female = - 5*0 + 5*1 - 4*0 + 4*1 - 1*0 + 1*1 = 10
Typical Male = - 5*1 + 5*0 - 4*1 + 4*0 - 1*1 + 1*0 = -10
Now, if we scored people who were clearly intersex, i.e. people with XX chromosomes, male genitalia and typical male secondary sexual characteristics, hormones and physiology or people with XY chromosomes, female genitalia and typical female secondary sexual characteristics, hormones and physiology we’d get:
Intersex (XX + male phenotype) = - 5*0 + 5*1 - 4*1 + 4*0 -1*1 + 1*0 = 0
Intersex (XY + female phenotype) = - 5*1 + 5*0 - 4*0 + 4*1 - 1*0 + 1*1 = 0
In addition to people who have the opposite phenotypic sex from their genotypic sex, intersex also includes people who’s phenotypic sex cannot be classified as either female or male. Their scores might look something like:
Intersex (XX + unclear phenotype) = - 5*0 + 5*1 - 4*0 + 4*0 -1*0 + 1*0 = 5
Intersex (XY + unclear phenotype) = - 5*1 + 5*0 - 4*0 + 4*0 - 1*0 + 1*0 = -5
Leonard Sax estimates the percentage of people clinically classified as intersex at 0.018%. Let’s assume that on average intersex people (as defined by Leonard) score 0 on our sex score.
But, there’s a little more complexity because a small percentage of the population has chromosomal abnormalities (i.e. something other than XX or XY). For instance, Klinefelter syndrome refers to people, typically classified as male, who have XXY chromosomes while Turner syndrome refers to people, typically classified as female, who have just one X chromosome.
If we wanted to account for these chromosomal abnormalities in our formula we could add an additional term to get the below Updated Sex Score. This new score would leave the previously discussed scores unchanged (changes in bold):
Updated Sex Score = - 5*(Has a Y chromosome) + 4*(Has no Y chromosome) - 4*(Has male genitalia) + 4*(Has female genitalia) - 1*(Has male secondary sexual characteristics, hormones and physiology) + 1*(Has female secondary sexual characteristics, hormones and physiology) + 1*(Has 2 X chromosomes)
Along with other symptoms, individuals with Klinefelter syndrome occasionally develop some female secondary sexual characteristics such as broad hips and enlarged breasts. Let’s assume an individual like that scores a 1 on (Has male secondary sexual characteristics, hormones and physiology) but also scores a 0.5 on (Has female secondary sexual characteristics, hormones and physiology). Someone like that would score as below:
Person with Klinefelter = - 5*1 + 5*0 - 4*1 + 4*0 - 1*1 + 1*0.5 + 1*1 - 1*0 = -8.5
*Disclaimer: I have no expertise on Klinefelter or any of these conditions. I’m simply using them to illustrate how we can think about the ranges of genotypic and phenotypic variance.
In addition to people who have obviously atypical phenotypes for their genotypic sex and those with chromosomal abnormalities, there are also people with conditions such as LOCAH which affects somewhere between 0.1% and 2% of the population, and can also lead to atypical phenotypic development.
For illustrative purposes (not accuracy), let’s assume that on average people with chromosomal abnormalities or LOCAH who have a Y chromosome score -8.5 while people with chromosomal abnormalities or LOCAH who have no Y chromosome score +8.5.
Some definitions of intersex, such as that from Anne Fausto-Sterling, include all of these conditions. An estimated 1.7% of the population would count as intersex under that definition.
Now, imagine that we sampled a million people, got their sex scores and then estimated a sex-score density function using the sample data. Depending how much smoothing we apply, we might get something like this:
Note: To create these plots I simulated sex score data for sub-populations of typical males, females, intersex and those with chromosomal abnormalities/LOCAH assuming the average scores for each population and estimated relative sizes of these sub-populations discussed above. If you’re interested you can access the Jupyter notebook here.
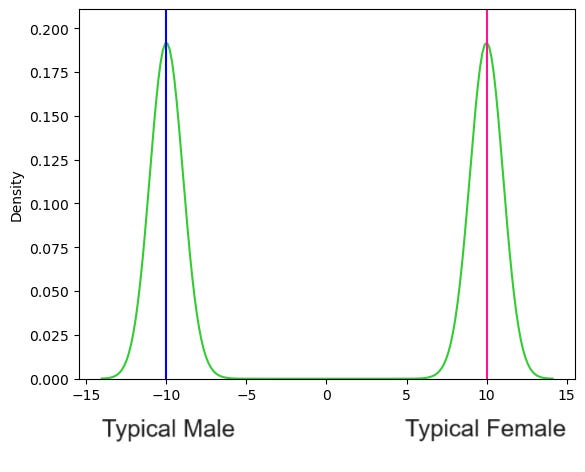
Sex looks like a bimodal distribution. We could then choose some boundaries to classify who is male and who is female. Given that the “Typical Male” scores a -10 while the “Typical Female” scores a +10, we might say that anyone with a score < -7.5 is categorized as male while anyone with a score > 7.5 is categorized as a female:
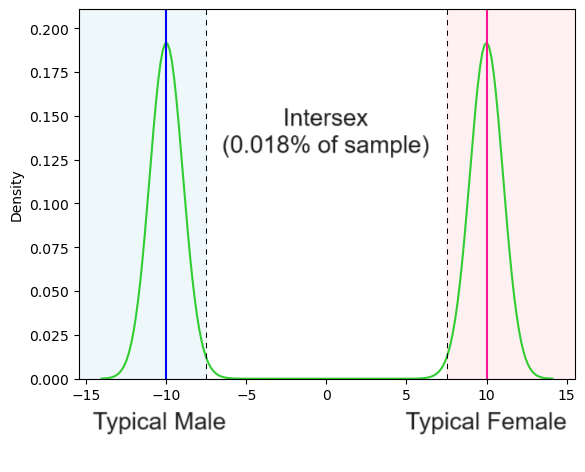
Given where we placed the boundary and how the data was simulated, this leaves us with only 0.018% of the population classified as intersex.
Now, if we estimated the density function with less smoothing we might see clusters representing some of the other conditions where people are not clearly intersex but are also not quite genotypically or phenotypically “normal”.
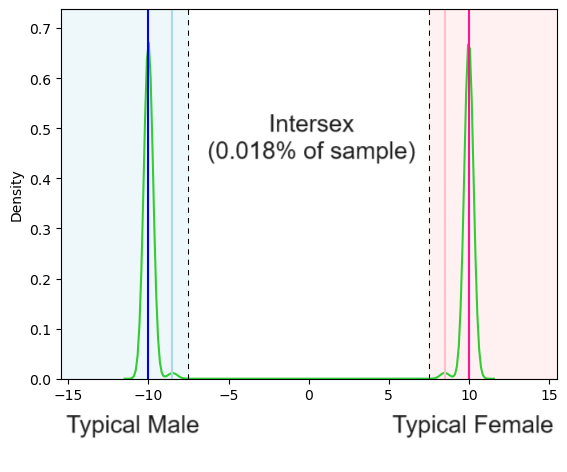
Now we can see small peaks in the density estimate to the right of the typical male score and to the left of the typical female score respectively reflecting the sub-populations with chromosomal abnormalities, LOCAH etc. And if we wanted to choose a boundary that matched up with the Anne Fausto-Sterling definition of intersex we could say anyone with a score below -9 is a man and anyone with a score above +9 is a woman which would result in a larger portion classified as intersex:
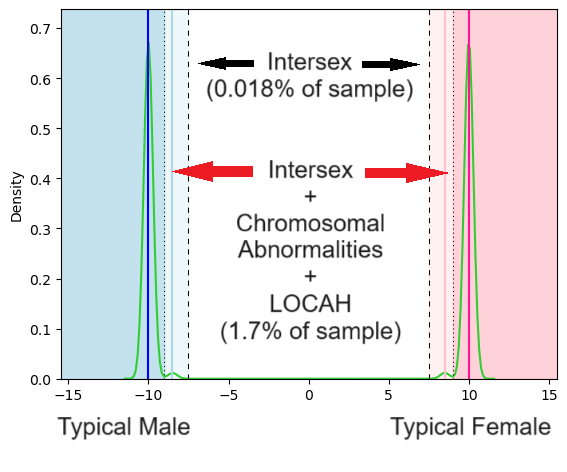
If you choose very tight boundaries around the “Typical Male” and “Typical Female” scores you get a lot more intersex people than you do if you choose slightly looser boundaries. But any reasonable boundary would capture most people as either male or female. And of course you could also choose to weight or measure the sex characteristics differently than I did in my made up score. All of those choices would impact who is or isn’t defined as intersex. In addition, the characteristics included in the score can added to or subtracted from. The most highly-weighted characteristics in my score were related to chromosomes given their centrality in clinical definitions, but chromosomes weren’t even discovered until at least the late 1800s. Despite that, people had a functional concept of sex, and one which would’ve almost entirely overlapped with our current concept.
To Summarize:
Sex - Evolutionary Biology Definition:
The biological definition of sex categorizes individuals based on the size of the gametes they produce. In humans and all complex sexually reproducing species organisms produce gametes of two distinct sizes.
Organisms that produce larger gametes are female, those that produce smaller gametes are male and those that produce both are hermaphrodites (either sequential or simultaneous). Organisms that don’t produce gametes are sexless.
Thus defined ,sexes are life-history stages that organisms pass through rather than identity characteristics. This definition of sex fails to classify many organisms that will eventually have a sex including juveniles.
Sex - Clinical Definition
The clinical definition of sex categorizes individuals based on the degree to which they share a set of sex-characteristics, both genomic and phenotypic.
Thus defined sex forms a strongly bimodal distribution. Individuals that do not fit the typical genomic-phenotypic patterns are classified as intersex and the percentage of people who are considered intersex depends on how loosely you define the sex category boundaries.
And to reiterate: just as there are some areas where gender is the relevant consideration and some where sex is, there are also some areas where the biological definition of sex is relevant (for humans) and some where the clinical definition is.
The level of interest around sex, how it’s defined and whether it’s binary, is related to the debate around sex differences and gender expression. While pretty much everyone can agree that there are substantial measured differences between the sexes in terms of strength and size and smaller measured differences in terms of psychological characteristics and interests, the cause of these differences is controversial. In my next post (or posts) I’ll review some of the evidence for and theory explaining sex differences in humans and how that relates to my understanding of gender and gender identity.
“Because no organism has infinite resources, organisms can either produce many small gametes, making it more likely that some of them will find a partner, or produce fewer but larger gametes, making it more likely that the new individual will have what it needs to survive and thrive. In models of the transition from isogamy to anisogamy, as mutations introduce differences in gamete size, two winning strategies emerge [...]. One is to produce a large number of small gametes – too small to create viable offspring unless they recombine with a larger, well-provisioned gamete. The other winning strategy is to produce a few, large, well-resourced gametes which can create viable offspring, no matter how small the other gamete they fuse with. Intermediate approaches, such as producing a moderate number of moderately well-provisioned gametes, or a mixture of differently sized gametes don’t do as well. Organisms that try to follow these ‘middle ways’ leave less offspring than their competitors and the population evolves to the two extremes - small gamete makers and large gamete makers.”
Paul E. Griffiths, What are biological sexes? p. 8, University of Sydney (2021)
“The Australian saltwater crocodile (Crocodylus porosus) lays eggs that develop into gigantic, highly territorial males if incubated between 30 and 33 degrees Celsius. At higher or lower temperatures genetically identical eggs develop into females. Like many other reptiles, the crocodile uses incubation temperature and not chromosomes to switch eggs into male or female pathways of development (Lang and Andrews 1994).”
Paul E. Griffiths, What are biological sexes? p. 13, University of Sydney (2021)
“Mating types exist when gametes are identical in size and structure, but the genome of each gamete contains genetic markers which affect which other gametes it can combine with. Typically, gametes with the same genetic marker can’t combine with one another and one function of mating types is to prevent genetically similar individuals from mating, which can have deleterious genetic effects. Some species have many hundreds of mating types, and newspapers often report research into this phenomenon under headlines like: ‘Scientists discover species with hundreds of sexes!’ But most biologists refer to these as ‘mating types’ and reserve the term ‘sexes’ for gametes that are different in size and structure [...].”
Paul E. Griffiths, What are biological sexes? p. 11, University of Sydney (2021)
“In some species several, distinct regions of phenotypic space have evolved as ways to pursue one gametic strategy. The Ruff, a common wading bird, has three strikingly different male forms (Figure 2). ‘Independent’ males are large birds with costly ornamental plumage which they display to attract mates. Independents defend small territories on the ‘lek’, the area where displays are made. ‘Faedermales’ are smaller and have the same plumage as females. They invest their energy in larger testicles and increased sperm production. ‘Satellitemales’ have a different colour of display plumage and do not defend territories. All three male morphs of the Ruff are pursuing a reproductive strategy focused on the production of large numbers of small gametes, but using different tactics to do so. So three distinct regions of Ruff phenotypic space are ways of being male.”
Paul E. Griffiths, What are biological sexes? p. 3, University of Sydney (2021)
The biologists' view of sex as a life stage is interesting because it gets at something fundamental about life being a process. One can contrast this with how humans, both individually and as a collective, view themselves as individuals that have an essence - in other words - indenties. Now sex happens to be an important concept (as are most concepts) that laid the foundation for god knows how many other breakthroughs. But there's something interesting about the way we take concepts that define differences between individuals and turn it into an identity. I'm guessing this is a natural function of being social mammals since most of our higher level faculties including language probably evolved to help us relate to one another. It therefore makes sense that concepts that differentiate us would be intergral to us - since the only way we can talk about ourselves is by comparing and contrasting with others. There's no real argument here, just a semi-stoned rant.
If the production of gametes determines biological sex, why isn't the preparation (i.e. childhood) part of that overall production process? The preparation seems like it would be very important biologically. Then after fertility, protecting one's offspring would also seem to be part of the overall gamete production process.
Therefore, if doctors have ways to pretty accurately predict which types of gametes an individual would produce as a fertile adult, then that seems like a way to determine biological sex for juveniles and non-fertile individuals.